2017.02.03.3
Files > Volume 2 > Vol 2 No 3 2017 > Investigaciones
INVESTIGATION / RESEARCH
Estudio comparativo in vitro de estrategias adaptativas en especies de Hylocereus, Cactaceae, con distribución ecológica contrastada
In vitro comparative study of adaptive strategies in Hylocereus species, Cactaceae, with contrasted ecological distribution
Máximo Moreira-Palacios1*, Aminael Sánchez-Rodríguez1
http://dx.doi.org/10.21931/RB/2017.02.03.3
_______________________________________________________________________________________________________________________
RESUMEN
Hylocereus ocamponis e Hylocereus triangularis son dos especies de cactáceas estrechamente relacionadas filogenéticamente pero que muestran hábitos de crecimiento completamente contrastados. La primera abunda en ecosistemas secos y la segunda en bosque tropical lluvioso, en bosques secos occidentales y la región amazónica del Ecuador, respectivamente. En el presente trabajo se empleó el cultivo in vitro como plataforma para el estudio de adaptaciones en ambas especies. El cultivo in vitro ofrece la posibilidad de estudiar de forma comparada la respuesta de explantes a reguladores del crecimiento en condiciones altamente controladas. Se evaluaron combinaciones de reguladores de crecimiento thidiazuron (TDZ), bencil amino purina (BAP), ácido naftalenacético (NAA), ácido 2,4-diclorofenoxiacético (2,4-D) y Kinetina (KIN) en diferentes tipos de explantes para estudiar sus respuestas morfogenéticas y hacer una relación con la tolerancia al estrés y capacidad adaptativa (plasticidad fenotípica) en H. ocamponis y H. triangularis. Los explantes de H. triangularis mostraron un mayor rango dinámico de respuesta a los tratamientos, especialmente durante la formación de cladodios y callos; la mejor formación de brotes (1,5 por explante) y callos (0,75 por explante) fue al aplicar 0,5 µl de TDZ con 0,5 µl de NAA. Los explantes de H. ocamponis mostraron casi siempre una inhibición ante los tratamientos y la mejor respuesta fue a la formación de raíces (1,43 por explante con 5 µl de BAP) lo que puede estar directamente relacionado con su hábitat de procedencia. El cultivo in vitro resultó ser una metodología útil para el estudio de adaptaciones en especies con distribución ecológica contrastada y reveló una gran plasticidad en H. triangularis lo que concuerda con su capacidad de expansión de hábitat.
Palabras clave: Caryophyllales, Cactaceae, Hylocereus ocamponis, Hylocereus triangularis, cultivo in vitro, estrés abiótico, reguladores de crecimiento vegetal, plasticidad fenotípica en plantas
_______________________________________________________________________________________________________________________
ABSTRACT
Hylocereus ocamponis and Hylocereus triangularis are two phylogenetically closely-related cacti species that show completely different growth habits. The first species occurs in dry ecosystems while the second one growths in the tropical rainforest, in dry forest western and the amazonic region Ecuadorian. In this work, in vitro culture was used as platform to study adaptations in both species. In vitro culture offers the possibility to compare responses to growth regulators under highly controlled conditions. Combinations of growth regulators (TDZ, BAP, NAA, 2,4-D and KIN) were evaluated in different types of explants to study their morphogenetic responses and make a connection with stress tolerance and adaptive capacity (phenotypic plasticity) in H. ocamponis and H. triangularis. H. triangularis showed a greater dynamic range of responses to the assayed treatments, especially during callus and cladode formation. Treatments produced in most cases an inhibition in H. ocamponis explants compared to the untreated control except during root induction, which can be directly related to its growing habits. In vitro culture proved to be a useful methodology to study adaptations in species with contrasting ecological distributions and revealed a great plasticity in H. triangularis that is consistent with its capacity to conquer new habitats.
Keywords: in vitro culture, stress, Hylocereus, growth regulators, plant phenotypic plasticity
_______________________________________________________________________________________________________________________
INTRODUCCIÓN
La relación entre distribución geográfica, hábitos de crecimiento, adaptabilidad, supervivencia y conservación son temas aún no comprendidos a cabalidad en varias especies vegetales.1-3 Una de las razones que podría explicar este vacío en nuestro conocimiento es la lenta tasa de crecimiento de muchas plantas,4,5 lo que impide estudiar sus adaptaciones in situ. Una alternativa para circunvalar esta limitación reside en estudiar las adaptaciones de especies de interés en condiciones in vitro. El cultivo in vitro ofrece la posibilidad de controlar y acelerar las condiciones de crecimiento en especies vegetales y se ha demostrado que es una herramienta que permite estudiar mecanismos adaptativos en plantas.6,7
El empleo de métodos de cultivo in vitro para acelerar el crecimiento vegetal es particularmente atractivo en cactáceas, las cuales son ampliamente conocidas por su lento desarrollo en condiciones naturales. El crecimiento in vitro de cactáceas se acelera debido a la alta humedad relativa y alta concentración de azúcares que facilitan un incremento en el rango fotosintético.8-10 Se ha comprobado que Coryphantha minima, una cactácea en peligro de extinción, creció 7 veces más rápido en condiciones in vitro que en su medio natural. 11
El cultivo in vitro se basa en el principio de la totipotencia celular, el cual establece que cualquier célula vegetal puede regenerar una planta completa.8 Las técnicas de cultivo in vitro permiten estudiar procesos fisiológicos y bioquímicos que se dan en las plantas como respuesta a condiciones ambientales, nutricionales u hormonales. Estos procesos son determinados por varios factores como el tipo y tamaño del material vegetal, constitución genética, edad del órgano o tejido, estado fisiológico/sanitario, ubicación del explante dentro de la planta e incluso la forma en la que se coloque el explante en el medio de cultivo. 8,12,13
Es importante señalar que el cultivo in vitro como tal constituye una fuente de estrés para el material vegetal el cual debe adaptarse a las nuevas condiciones in vitro. El estrés se define como la exposición a condiciones ambientales potencialmente adversas (estresores) que alteran el crecimiento y desarrollo de una planta desencadenando una amplia gama de respuestas, que pueden ir desde una alteración de su expresión génica y del metabolismo celular a cambios en su crecimiento. 14,15 Superada la fase de introducción in vitro, el material vegetal se expone a nuevas condiciones de desarrollo y por tanto de estrés, siendo una de las más comunes el uso de reguladores de crecimiento vegetal (auxinas, citoquininas, giberelinas, entre otros). Los reguladores de crecimiento promueven la diferenciación y desdiferenciación de tejidos mediante variados mecanismos entre los cuales está el control del ciclo celular, y además son conocidos por generar una diversidad de respuestas.12 Durante el cultivo in vitro se pueden observar otros fenómenos no deseados como la hiperhidricidad que es inducida principalmente por las citoquininas 16,17 o procesos de fenolización, desarrollo radicular anormal e ineficiente o mal formación de brotes como respuesta al estrés.18,19 La respuesta observada dependerá en gran medida del tipo de explante los cuales varían significativamente entre sí.
Aunque en principio cualquier tejido u órgano de la planta puede ser utilizado como explante de inicio para la introducción in vitro, de éste, en gran parte, va a depender el proceso de diferenciación y propagación y por ende la calidad de sus propágulos. En el caso de las Cactáceas, dada su particularidad de formas, se emplean explantes que contengan areolas, con cortes en partes basales, medias o apicales en las cuales se evalúan condiciones óptimas para activar respuestas morfogenéticas.20,21 A partir de estos explantes, el cultivo in vitro en cactáceas se ha utilizado como una alternativa a la propagación tradicional por semillas ya que estas pueden presentar bajos rangos de germinación.13 El comportamiento que tienen las cactáceas frente a los reguladores de crecimiento es particularmente variable entre especies. Varios autores sostienen que estas diferencias no son debidas a variaciones en los niveles endógenos hormonales entre especies, ya que las cactáceas de forma general presentan altos niveles endógenos de auxinas y de citoquininas. 22-26 Esto sugiere que la variabilidad de respuesta a condiciones in vitro entre especies se deba fundamentalmente a otros factores genéticos, no relacionados con la expresión de genes que participan en la síntesis de hormonas endógenas, y epigenéticos. Estos últimos son indicadores directos de diferencias en adaptaciones a niveles de estrés entre especies. 27
Se conoce que las respuestas de las especies vegetales a las condiciones de cultivo in vitro dependen de su genotipo.22 Las células vegetales que entran al proceso de desdiferenciación in vitro, borran una parte de sus marcas epigenéticas (modificaciones del ADN como la metilación que determinan el estado de expresión de varios genes) para recuperar su totipotencia.28 Es por ello que las diferencias en la capacidad regenerativa entre genotipos pudiera deberse a la rapidez o facilidad con que las marcas epigenéticas se borran o reprograman.29 Adicionalmente las plantas pueden tener una memoria de estrés en su cromatina, probablemente característica de las condiciones que enfrentan en su hábitat natural y que influye en las reacciones de la célula y su desarrollo. 18,30
En el presente trabajo se estudiaron adaptaciones en dos especies de cactáceas del género Hylocereus las cuales se encuentran naturalmente distribuidas en dos regiones tropicales y subtropicales.31 La particularidad de estas especies es que a pesar de su cercanía genética difieren considerablemente en su hábitat natural. Mientras que H. triangularis se encuentra en bosques húmedos de la amazonía con precipitaciones sobre los 2000 mm anuales,32 H. ocamponis se encuentra en bosques secos que no sobrepasan los 600 mm anuales.33 Morfológicamente H. ocamponis es más globosa que H. triangularis, y es similar a otras especies de hábitats cálidos y secos que presentan mayor globosidad.34 Aunque no se han encontrado reportes que comparen la generación de raíces en estas dos especies, este puede ser distinto dado que una especie vive en un ambiente hidromórfico y la otra en un xeromórfico.35
El conocimiento de la capacidad adaptativa y formas de desarrollo en base a su hábitat natural en especies de Hylocereus permitirían plantear estrategias de manejo y conservación de estas especies. Ello es particularmente importante en el caso de H. ocamponis que se encuentra con un alto nivel de riesgo en sus poblaciones.36 Para generar conocimiento sobre el potencial adaptativo y plasticidad en H. ocamponis y H. triangularis se utilizó el cultivo in vitro como plataforma de estudio ya que permite someter a estas especies ante los mismos tipos de estrés (reguladores del crecimiento) en un ambiente altamente controlado. La hipótesis de trabajo fue que la diferencia entre las respuestas de los explantes de ambas especies ante un mismo estrés será proporcional a su capacidad adaptativa para acomodar dicho estrés. Es conocido que para tolerar el estrés las plantas activan mecanismos fisiológicos que les permitan, bajo esas condiciones, adaptarse o morir.37,38 El éxito de estos mecanismos podría depender de su genotipo, de su plasticidad epigenética y de la historia evolutiva que han seguido para adaptarse al hábitat del cual provienen. Considerando la cercanía filogenética de las dos especies estudiadas,39,40 lo cual impone una similitud en su genotipo y por ende en su plasticidad epigenética, las diferencias en sus respuestas al estrés in vitro podrían relacionarse de forma directa a adaptaciones en sus hábitats contrastados.
METODOLOGÍA
Desinfección y siembra de semillas
Semillas de H. triangularis e H. ocamponis se introdujeron en condiciones in vitro para obtener el material de partida. La importancia de trabajar con semillas es porque toleran los tratamientos de desinfección a diferencia de los cladodios de cactáceas que en sus areolas tienen gran cantidad de esporas de hongos.41-44 Las semillas se desinfectaron con 200 ml de agua destilada esterilizada con cinco gotas de jabón, seguido de etanol al 70% por 30 segundos y cloro al 1% por 10min, cada paso fue alternado con enjuagues de agua destilada esterilizada. Una vez sembradas en medio semisólido con 2,5g/L de nitrofoska foliar, con 7 g/l de agar, 20 g de sacarosa y pH de 5.80 ± 0.02, se mantuvieron a 25ºC con fotoperiodo de 12 horas luz y 12 horas oscuridad, hasta obtener plántulas mayores a 2 cm de largo y de ellas obtener explantos.
Obtención y siembra de explantes
Previas separaciones de las raíces formadas de cada plántula de dos centímetros se obtuvieron explantos de 0.5 cm, considerando cuatro tipos de explante; apical (A), medio apical (MA), medio basal (MB) y basal (B). En plántulas mayores a dos centímetros se aplicó el mismo proceso de división en 4 partes. Cada tipo de explante fue sembrado en medio con reguladores de crecimiento y el testigo. El control fue un medio Murashige & Skoog (1962), MS sin reguladores de crecimiento y con cada uno de los tipos de explante.
Para todos los tratamientos de desarrollo in vitro de los cladodios se utilizó el medio MS 45 semisólido con 7 g/l de agar, 20 g de azúcar y pH 5.80 ± 0.02 ajustado con soluciones de NaOH 1N o HCl 1N, y autoclavado a una presión de 1 kg/cm2 y temperatura de 120ºC por 20 minutos. En cada frasco se puso aproximadamente 25 ml de medio. A este medio se le adicionaron diferentes reguladores de crecimiento constituyendo los tratamientos aplicados. Se utilizaron 2 citoquininas Thidiazuron, TDZ (0,1 y 0,5 µM) y Bencil amino purina, BAP (1 y 5 µM) solas o combinadas con ácido naftalenacético, NAA 0,5 µM,46-48 y como medio inductor de callos se utilizó ácido 2,4-diclorofenoxiacético, 2,4 D 9 µM + Kinetina, KIN 4,6 µM.49 Ver en la tabla 1 todos los tratamientos aplicados. Los ensayos se mantuvieron a 25ºC con fotoperiodo de 12 horas luz y 12 horas oscuridad.
Diseño y análisis de datos
De cada tratamiento se hicieron 2 repeticiones en cada una de las especies de Hylocereus. Una repetición consta de 5 frascos. En cada frasco 4 explantes de 0.5 cm (sub-muestras). Los datos de brotación, enraizamiento y formación de callos se tomaron a los 45 días. Se consideraron el porcentaje de explantes que responden por tratamiento y tipo de explante utilizado (apical, medio apical, medio basal y basal). Los explantes se asignaron a cada tratamiento de forma completamente aleatoria y así mismo los tratamientos se realizaron con un diseño completamente aleatorio.
Para representar la formación de cladodios, raíces y callos se utilizaron plots del programa R GNUR versión R-3.2.1.50 Cada barra simboliza el promedio de cada respuesta (40 réplicas), a la cual se restó el promedio del tratamiento control de su especie correspondiente (40 réplicas). Con ello se obtuvo la desviación de cada tratamiento, el cual indica el efecto que tiene el tratamiento sobre cada respuesta. El tratamiento control al formar naturalmente brotes y raíces sirvió como modelo para evaluar el estrés al que están sometidos los explantos expuestos a diferentes combinaciones de reguladores de crecimiento.
RESULTADOS
En ambas especies los cladodios se obtuvieron a partir de plántulas desarrolladas de semillas. El tratamiento de desinfección fue exitoso ya que la contaminación no superó el 5% en ninguna de las dos especies. Tanto H. ocamponis como H. triangularis presentaron alta variación en la capacidad de respuesta de sus explantes para formar brotes, raíces y/o callos. Solo en tres de los diez tratamientos ensayados en H. ocamponis se logró una respuesta que alcanzara el 80% de sus explantes. En el caso de H. triangularis esto ocurrió en seis tratamientos (Tabla 1).
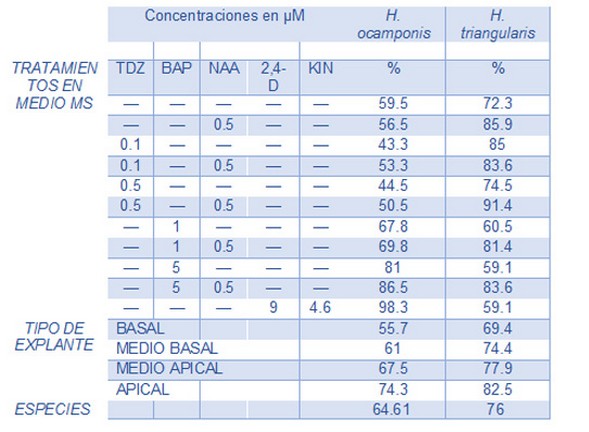
Tabla 1. Porcentaje de respuestas presentadas según el Tratamiento, Tipo de Explante y la Especie, a los 45 días de experimentación. La tabla indica el porcentaje de explantes que presentó algún tipo de respuesta (cladodio, raíz o callo) en función del tratamiento, el tipo de explante y de forma general la especie de origen. Cada fila indica el tratamiento utilizado, en la cual puede estar el regulador de crecimiento vegetal solo o combinado con otro. El guion representa la ausencia de un regulador.
Los explantes de H. triangularis responden de manera general en mayor proporción que los de H. ocamponis. El necrosamiento fue mayor en H. ocamponis.
RESPUESTA IN VITRO SEGÚN EL ORIGEN DEL EXPLANTE
En ambas especies de Hylocereus el explante apical fue el que mejor respondió frente a todos los tratamientos (Tabla 1). La formación de raíces presentada en el explante apical fue significativamente mejor que la del resto de explantes, mientras que en formación de cladodios (brotes) y callos no se observaron diferencias significativas. Todos los resultados que se presentan a partir de esta sección se refieren a la respuesta de explantes apicales para ambas especies.
FORMACIÓN DE RAÍCES POR EXPLANTO EN LAS ESPECIES ESTUDIADAS
En la mayoría de las respuestas a los tratamientos se observó una inhibición en formación de raíces a excepción de los tratamientos con 5 µM BAP sólo o combinado con 0.5 µM NAA en H. ocamponis, y de los tratamientos con 0.5 µM NAA, 0.1 µM TDZ sólo o combinado con 0,5 µM NAA en H. triangularis. Al usar 5 µM BAP sólo se obtuvo una media de 1,43 raíces por explanto y combinado una media de 1,36 ambas con p<0,001 en H. ocamponis, mientras que en H. triangularis al aplicar 0.5 µM NAA se obtuvo una media de raíces por explante de 1,26 con p<0,012. El tratamiento con 5 µM BAP provocó la mayor diferencia en cuanto a la formación de raíces entre las dos especies (Figura 1a). Ello se debe a que en H. ocamponis fue el tratamiento que más enraizamiento provocó, mientras que en H. triangularis fue uno de los que ocasionó mayor inhibición en formación de raíces respecto al control. De igual manera la respuesta al tratamiento con 0.5 µM NAA mostró una alta diferencia entre las dos especies, pero en este caso de una forma contraria: formación en H. triangularis (Figura 1b) e inhibición en H. ocamponis con relación a sus respectivos controles.
En H. ocamponis el BAP en alta concentración influyó mejor que el TDZ sobre la formación de raíces. En H. triangularis bajas concentraciones de TDZ o BAP solas y combinadas generan mayor cantidad de raíces que altas concentraciones de los mismos reguladores. La mayor formación de raíces en H. triangularis se logró con el tratamiento de 0.5 µM NAA.
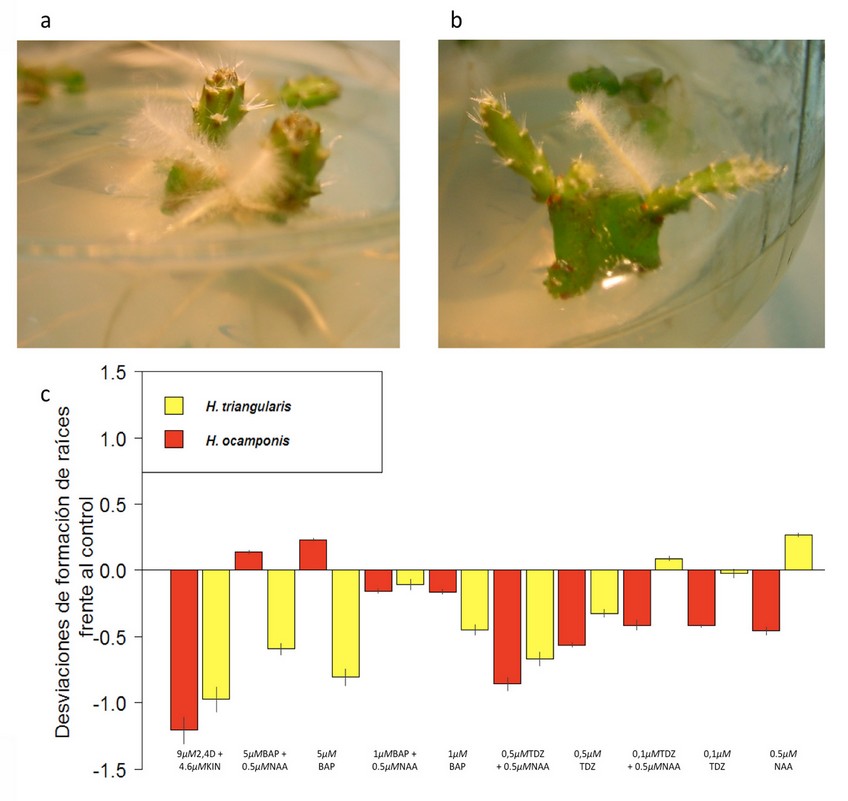
Figura 1. Formación de raíces por explanto in vitro en las dos especies de Hylocereus. a. Enraizamiento de H. ocamponis con el tratamiento 5 µM BAP. b. Enraizamiento de H. triangularis con el tratamiento 0.5 µM NAA. c. Desviaciones de la formación de raíces frente al control. La amplitud de las barras en el eje Y representa la desviación en el número de raíces formadas de ambas especies respecto a sus respectivos controles. Las desviaciones se obtuvieron al restar el valor medio de raíces por explante del control del valor medio de raíces por explante de cada tratamiento (eje X). La media del número de raíces por explanto del control fue de 0.98±0.09 y 1.21±0.11, en H. triangularis y H. ocamponis, respectivamente. Los errores estándar se muestran en función de las mediciones de los tratamientos sobre cada barra. El tratamiento con 9 µM 2,4D + 4.6 µM KIN no presenta error estándar puesto que no hubo ninguna respuesta de enraizamiento en ambas especies, por tanto, las barras representan el valor de los controles en el sentido negativo.
En ambas especies se observaron conjuntos de pelillos radicales, los cuales estaban distribuidos a lo largo de las raicillas. La formación de pelillos fue mayor en H. ocamponis, siendo el control (Figura 2) el que presentó mayor formación seguido de los tratamientos con BAP. Mientras que en H. triangularis se observaron en los tratamientos control y con 0.1 µM TDZ.

Figura 2. Formación de pelillos radicales en los tratamientos control de ambas especies de Hylocereus a. H. ocamponis b. H. triangularis.
La longitud de las raíces en función de los tratamientos (datos no presentados) mostró de forma general el mismo patrón que el de formación de raíces en ambas especies. Sin embargo, en H. ocamponis el tratamiento con 5 µM BAP+ 0.5 µM NAA tuvo mayor crecimiento de sus raíces, en contraste con el tratamiento de 5 µM BAP que fue la que mayor formación de raíces indujo en H. ocamponis. De igual manera, se observó que el BAP genera mayor crecimiento en las raíces de H. ocamponis que el TDZ, mientras que en H. triangularis el TDZ genera mayor crecimiento que el BAP. Sin embargo, en H. triangularis, se observó que altas concentraciones de TDZ influyen negativamente sobre el crecimiento de sus raíces y si está combinado con NAA disminuye aún más su crecimiento.
FORMACIÓN DE CALLOS A PARTIR DE EXPLANTES EN LAS ESPECIES ESTUDIADAS
No todos los tratamientos estimularon la formación de callos en las especies estudiadas. Sin embargo, tal y como se esperaba, la mayor inducción de callos en las dos especies fue con 9 µM 2,4D + 4.6 µM KIN, con 0,64 y 0,51 callos por explantes en H. triangularis e H. ocamponis, respectivamente y en ambos casos con p<0,001. Sin embargo, los callos se observaron de forma general en mayor cantidad en H. triangularis. El tratamiento que indujo la respuesta con mayor contraste para la formación de callos entre las especies fue el de 0.5 µM TDZ + 0.5 µM NAA. Al igual que en el crecimiento de raíces, los tratamientos con 5 µM BAP y 5 µM BAP + 0.5 µM NAA también indujeron respuestas contrastadas (Figura 3).
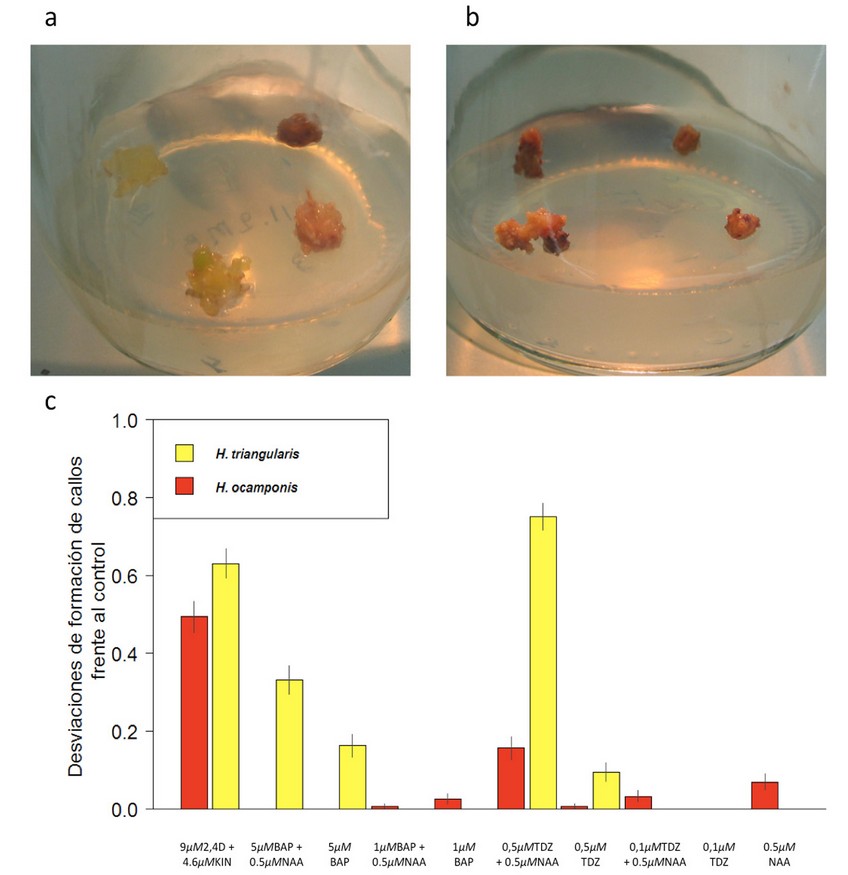
Figura 3. Formación de callos in vitro en las dos especies de Hylocereus. a. Callos formados en H. ocamponis con el tratamiento 9 µM 2,4D + 4.6 µM KIN. b. Callos formados en H. triangularis con el tratamiento 9 µM 2,4D + 4.6 µM KIN. c. Desviaciones de la formación de callos de ambas especies frente al control. La amplitud de las barras en el eje Y representa la desviación en el número de callos formados de ambas especies respecto a sus respectivos controles. Las desviaciones se obtuvieron al restar el valor medio de los explantos transformados en callos del control del valor medio de los explantos transformados en callos de cada tratamiento (eje X). La media de formación de callos del control fue de 0 en ambas especies. Los errores estándar se muestran en función de las mediciones de los tratamientos sobre cada barra. La ausencia de barras es porque no hubo formación de callos en ese tratamiento. Dado que el tratamiento control no presentó callos todos los tratamientos provocan una respuesta nula o en el sentido positivo, pero nunca en el sentido negativo del eje Y.
La mayoría de callos en ambas especies fueron de aspecto transparente, sin embargo, se observaron otros de color verde amarillento los cuales podrían ser más viables.51 En pocos casos se observaron explantos con aparente hiperhidricidad. La mayoría de callos tenían apariencia granular no friable, es decir no se desmenuza fácilmente, similar a lo observado en Ananas.52
FORMACIÓN DE CLADODIOS O BROTES A PARTIR DE EXPLANTES EN LAS ESPECIES ESTUDIADAS
La mayoría de tratamientos tuvo un efecto inhibitorio para la formación de cladodios. Aquí es importante resaltar que para aquellos tratamientos que tuvieron un efecto estimulador, dicho efecto fue mucho mayor para H. triangularis presentando mayor cantidad de cladodios (Figura 4). Al usar 0.5 µM TDZ + 0.5 µM NAA se obtuvo una media de 1,5 cladodios por explante con p < 0,001 en H. triangularis; mientras que en H. ocamponis la mejor respuesta fue 5 µM BAP con una media de 0,48 cladodios por explante con p < 0,001. El tratamiento que presentó el mayor contraste fue 0.5 µM TDZ + 0.5 µM NAA, en este tratamiento se presentó mayor cantidad de macizos de proliferación de brotes.
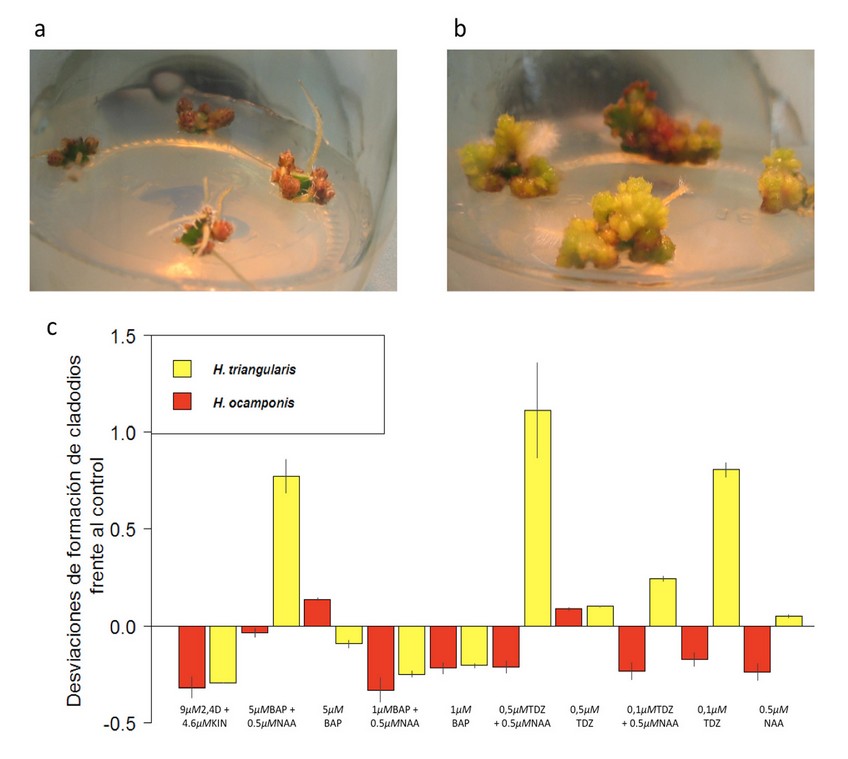
Figura 4. Formación de cladodios por explante in vitro en las dos especies de Hylocereus. a. Formación de cladodios de H. ocamponis con el tratamiento 5 µM BAP. b. Formación de cladodios de H. triangularis con el tratamiento 0.5 µM TDZ + 0.5 µM NAA. c. Desviaciones de la formación de cladodios frente al control. La amplitud de las barras en el eje Y representa la desviación en el número de cladodios formados de ambas especies respecto a sus respectivos controles. Las desviaciones se obtuvieron al restar el valor medio de cladodios formados por explante del control del valor medio de cladodios formados por explante de cada tratamiento (eje X). La media del número de cladodios por explante del control fue de 0.41±0.07 y 0.35±0.07, en H. triangularis y H. ocamponis, respectivamente. Los errores estándar se muestran en función de las mediciones de los tratamientos sobre cada barra.
Algunos explantos tomaron una coloración marrón (Figura 5a), la cual se debe a la oxidación de compuestos fenólicos en las áreas donde se cortaron los explantos y no afecta su producción de brotes.53 En H. ocamponis se observaron brotes de color rosado (Figura 5b), lo cual también ha sido reportado en otras cactáceas,54 aparentemente podrían ser betacianinas.55 También se observó que un alto número de explantos de ambas especies al ser cortados formaron una capa protectora que cubría el sitio donde se realizó el corte. Ésta capa es un polisacárido particular viscoso, el cual ha sido observado en otras especies en condiciones naturales, considerándolo como una adaptación de los cactus para colmatar y sellar rápidamente los tejidos dañados y evitar la pérdida de agua por evaporación, y de esta manera permite sobrevivir a la planta por largos periodos de tiempo.56

Figura 5. Cambios de coloración en explantos durante el proceso de brotación. a. Coloración marrón en H. triangularis con 5 µM BAP+NAA. b. Coloración rosada del nuevo cladodio de H. ocamponis, tratamiento control.
DISCUSIÓN
De manera general se observa un alto contraste en las respuestas de inducción de estructuras (raíces, callos y brotes) con respecto al control de cada especie, observándose que H. triangularis tiene más respuestas positivas que H. ocamponis. Los resultados contrastantes observados en la respuesta in vitro entre H. triangularis y H. ocamponis pudieran deberse a sus adaptaciones ecológicas, las cuales también son contrastantes ya que H. ocamponis proviene de bosque seco y H. triangularis de bosque húmedo.
La concentración 5 µM BAP es quizás la que mayor número de respuestas contrastantes provoca entre las dos especies para todas las estructuras analizadas. En H. triangularis la respuesta es positiva para la formación de callos, es decir tiene mayor inducción de esa estructura respecto a su control. En H. ocamponis en cambio la respuesta es positiva para la formación de brotes y crecimiento de raíces. El BAP incrementa el número de replicaciones durante la fase S del ciclo celular y por ello tiene un buen efecto sobre la brotación y elongación, pero puede inhibir la metilación del ADN.57-59 La metilación puede provocar alteraciones en la transcripción genética sin alterar la secuencia del ADN (modificaciones epigenéticas) y es un mecanismo responsable de la plasticidad fenotípica. Es bien conocido que el grado de metilación del ADN está relacionado con la respuesta de las plantas al estrés y funciona como mecanismo de protección del ADN.27 El hecho de que el BAP produzca una respuesta contrastada en ambas especies unido a su relación con los niveles de metilación de ADN, confirma el hecho de que los niveles de estrés a los cuales H. ocamponis y H. triangularis están expuestas son bien diferentes.
Otro de los contrastes en respuesta in vitro se evidenció frente al tratamiento con 0.1 µM TDZ + 0.5 µM NAA. Este tratamiento produce una respuesta positiva en H. triangularis al formar cladodios y raíces, mientras que inhibe la formación de estas estructuras respecto al control en H. ocamponis. El TDZ es un potente generador de estrés en las plantas y las obliga a modificar procesos metabólicos para poder sobrevivir.60-62 Esa adaptación está determinada por la capacidad de las células de reiniciar su programa genético y epigenético con el fin de soportar el ambiente hormonal.28 La forma de actuar del TDZ en los procesos de regeneración es promoviendo la acumulación de iones minerales, 37 y de esta manera lo predispone al tejido a adaptarse a las condiciones en las que se encuentra. Se sabe además que el TDZ es un inductor de variabilidad genética,63 y que a pesar de ser producto sintético, pudiera ser beneficioso, en condiciones controladas, para H. triangularis que es una especie con tendencia a expandir su hábitat. H. ocamponis por el contrario es una especie que vive al límite, que presenta una serie de adaptaciones que le permiten sobrevivir en un ambiente mucho más adverso que el de H. triangularis y para la cual pudiera ser contraproducente generar individuos con tal variabilidad que pusiera en riesgo su adaptación.
H. triangularis presentó mejores resultados de formación de callos y brotes mostrando mayor plasticidad y adaptabilidad a las condiciones in vitro que H. ocamponis. Estos resultados son consistentes con la mayor capacidad explorativa y tendencia a extensión de su hábitat que se ha visto en H. triangularis. Esa mayor capacidad explorativa de H. triangularis se puede relacionar con su predisposición para el aumento de variabilidad genética lo cual se alinea con la propensión observada en esta especie para la formación de callos. Se sabe que los callos se caracterizan por tener mayor variabilidad genética.8,64 La rápida y mayoritaria formación de brotes también es un indicador de la capacidad de adaptación y plasticidad de H. triangularis. La formación de brotes es indispensable para la colonización de ecosistemas, comportamiento natural en poblaciones naturales de H. triangularis.65 Además, las respuestas de especies de un mismo género en condiciones in vitro pueden variar como un reflejo de su normal rango de crecimiento en sus condiciones naturales.66
Los procesos de diferenciación son particulares de cada especie y pueden depender más del hábitat del que provienen que del balance de auxinas y citoquininas,67 es más, el hábitat afecta los procesos de biosíntesis, metabolismo y transporte de auxinas y citoquininas.68-70 Es así que las condiciones ambientales del hábitat pueden conducir a una mayor flexibilidad genómica incluso en las generaciones sucesivas, y pueden aumentar el potencial de adaptación de las especies,71 morfológica, fisiológica y bioquímicamente.6,72 Un ejemplo de esas adaptaciones se da en algunas MAC (especies que tienen el metabolismo ácido de las crasuláceas), las cuales a su vez pueden ser MAC facultativas, es decir que pueden alternar con un tipo de fotosíntesis C3 cuando las condiciones de humedad son favorables.73 Esto se cumple en H. triangularis 74 y a su vez explica su mejor adaptación y crecimiento en condiciones in vitro que H. ocamponis.
La formación de raíces fue mayor en H. ocamponis, lo cual demuestra una mayor especialización, ya que es un comportamiento típico de especies de ecosistemas secos que están programadas genéticamente para formar una mayor masa radicular como estrategia de sobrevivencia.56,75 Esos procesos de enraizamiento también dependen de otros factores como reguladores de crecimiento, luz, temperatura,76,77 o el estrés hídrico, que reduce el contenido de citoquininas en la planta.78 Sin embargo es de considerar que en ambientes altamente estresantes la plasticidad de las especies es reducida.79 Esta reducción se explica porque en ambientes naturales el estrés induce procesos selectivos de combinaciones de rasgos que confieren mayor aptitud a ese medio a costo de disminución de la variación genética en las generaciones, llegando al punto de que las especies podrían perder la capacidad de adaptarse a diferentes condiciones ambientales.14 Esta reducción de la plasticidad podría estar presente en H. ocamponis y se refleja en su baja adaptabilidad in vitro frente a todos los tratamientos aplicados.
Una limitante del presente estudio es que no se analizaron los niveles endógenos de reguladores de crecimiento de cada una de las especies. Estos niveles endógenos varían en función de la especie.37 Las cactáceas son conocidas por la producción de niveles elevados de auxinas.26 Esta alta producción de auxinas es la explicación de que algunas cactáceas, como Mammillaria gracillis, sean capaces de formar callos en condiciones in vitro sin necesidad de reguladores de crecimiento.80 Considerando que la formación de callos se produce con hormonas solas o combinando auxinas y citoquininas, podríamos asumir que la concentración endógena de citoquininas también es elevada. Los elevados niveles endógenos de reguladores de crecimiento explicarían como bajos niveles de auxinas o de citoquininas exógenos incrementan la producción de brotes axilares y raíces en H. triangularis.25,81 Las respuestas contrastadas que se decriben en el presente trabajo frente a las mismas concentraciones exógenas de reguladores de crecimiento en ambas especies pudiera bien deberse a diferencias en sus niveles endógenos. Sin embargo, este supuesto no invalida las conclusiones a las que hemos podido llegar sobre sus procesos adaptativos.
Aunque el papel de los reguladores de crecimiento depende de muchos factores como su concentración, tipo entre otros, es posible que las diferencias en las poblaciones tanto en la producción como en la sensibilidad a diferentes hormonas vegetales pueda contribuir a generar los diferentes patrones de la respuesta plástica en las plantas, por lo que el estudio de poblaciones ecológicamente distintas puede ayudar a proveer información invaluable acerca de los mecanismos más próximos de sus respuestas plásticas.82 Por tanto la plasticidad observada en H. triangularis versus la especialización observada en H. ocamponis nos deja ver que H. triangularis al tener mayor plasticidad adaptativa puede ser más propensa a sobrevivir en condiciones ambientales nuevas y que sus poblaciones no estén comprometidas, esto es corroborado por 83 en su análisis de plasticidad fenotípica en plantas. Mientras que H. ocamponis al tener mayor especialización, sus poblaciones podrían estar comprometidas.
H. triangularis mostró mayor capacidad de respuesta frente a todas las condiciones de estrés in vitro que H. ocamponis. Nuestros resultados resaltan el hecho de que las poblaciones naturales de H. triangularis pudieran tener una mayor capacidad expansiva y ser más tolerante a cambios ambientales. H. ocamponis al presentar poca capacidad de respuesta in vitro nos lleva a pensar en que sus poblaciones pueden estar comprometidas en cuanto a su supervivencia ante cambios ambientales. El cultivo in vitro resultó ser de gran utilidad para evaluar procesos adaptativos en las especies estudiadas.
REFERENCIAS BIBLIOGRÁFICAS
1. López Soto JL, Ruiz Corral JA, Sánchez González J de J, Lépiz Idefonxo R. Adaptación climática de 25 especies de frijol silvestre ( Phaseolus spp ) en la República Mexicana. Rev Fitotec Mex. 2005;28: 221–230.
2. Bayuelo-Jimenez JS, Craig R, Lynch JP. Salinity tolerance of Phaseolus species during germination and early seedling growth. Crop Sci. 2002;42: 1584–1594.
3. Kadam NN, Yin X, Bindraban PS, Struik PC, Jagadish KS V, Rice I, et al. Does morphological and anatomical plasticity during the vegetative stage make wheat more tolerant of water deficit stress than rice? Plant Physiol. 2015;167: 1389–1401. doi:10.1104/pp.114.253328
4. Sun Y, Shen Y, Li A, Fu W, Wang Y, Ying Q, et al. Ectopic expression of Dendrobium EREB5 gene in Arabidopsis influences leaf morphology. Vitr Cell Dev Biol - Plant. 2014;50: 425–435. doi:10.1007/s11627-014-9604-6
5. Dhar PC, Awal MA, Sultan MS, Rana MM, Sarker A. Interspecific competition, growth and productivity of maize and pea in intercropping mixture. Sci J Crop Sci. 2013;2: 136–143. doi:10.14196/sjcs.v2i10.974
6. Cori P Di, Lucioli S, Frattarelli A, Nota P, Tel-Or E, Benyamini E, et al. Characterization of the response of in vitro cultured Myrtus communis L. plants to high concentrations of NaCl. Plant Physiol Biochem. 2013;73: 420–426.
7. Boamponsem GA, Leung DWM. Use of compact and friable callus cultures to study adaptive morphological and biochemical responses of potato (Solanum tuberosum) to iron supply. Sci Hortic (Amsterdam). Elsevier B.V.; 2017;219: 161–172. doi:10.1016/j.scienta.2017.03.012
8. George E, Hall M, De Klerk G-J. Plant Propagation by Tissue Culture 3rd Edition. Vol 1. The Background. Springer. 2008.
9. Malda G, Backhaus RA, Martin C. Alterations in growth and crassulacean acid metabolism (CAM) activity of in vitro cultured cactus. Plant Cell Tissue Organ Cult. 1999;58: 1–9.
10. Elias-Rocha M, Santos-Díaz M, Arredondo-Gómez A. Propagation of Mammillaria candida (Cactaceae) by tissue culture techniques. Haseltonia. 1998;6: 96–101.
11. Malda G, Suzán H, Backhaus R. In vitro culture as a potential method for the conservation of endangered plants possessing crassulacean acid metabolism. Sci Hortic (Amsterdam). 1999;81: 71–87.
12. Farhadi N, Panahandeh J, Azar AM, Salte SA. Effects of explant type , growth regulators and light intensity on callus induction and plant regeneration in four ecotypes of Persian shallot (Allium hirtifolium). Sci Hortic (Amsterdam). Elsevier B.V.; 2017;218: 80–86. doi:10.1016/j.scienta.2016.11.056
13. Pérez-Molphe-Balch E, Santos-Diaz MDS. Tissue culture of ornamental cacti. Sci Agric. 2015;72: 471–563. doi:10.1590/0103-9016-2015-0012
14. Grativol C, Hemerly AS, Ferreira PCG. Genetic and epigenetic regulation of stress responses in natural plant populations. Biochim Biophys Acta. 2011;1819: 176–85. doi:10.1016/j.bbagrm.2011.08.010
15. Kacperska A. Sensor types in signal transduction pathways in plant cells responding to abiotic stressors: do they depend on stress intensity? Physiol Plant. 2004;122: 159–168. doi:10.1111/j.0031-9317.2004.00388.x
16. Kataeva N, Alexandrova I, Butenko R, Dragavtceva E. Effect of applied and internal hormones on vitrification and apical necrosis of different plants cultured in vitro. Plant Cell Tissue Organ Cult. 1991;27: 149–154.
17. Vasconcelos A, Tomas LF, Camara TR, Willadino L. Hiperidricidade: uma desordem metabólica. Cienc Rural. 2012;42: 837–844.
18. Ruffoni B, Savona M. Physiological and biochemical analysis of growth abnormalities associated with plant tissue culture. Hortic Environ Biotechnol. 2013;54: 191–205. doi:10.1007/s13580-013-0009-y
19. Cassells AC, Curry RF. Oxidative stress and physiological, epigenetic and genetic variability in plant tissue culture: implications for micropropagators and genetic engineers. Plant Cell Tissue Organ Cult. 2001;64: 145–157.
20. Dabekaussen M, Pierik R, Van der Laken J, Hoek Spaans J. Factors affecting the areole activation in vitro in the cactus Sulcorebutia alba Raush. Sci Hortic (Amsterdam). 1991;46: 283–294.
21. Gomes FLAF, Heredia FF, Silva PBE, Facó O, Campos FDADP. Somatic embryogenesis and plant regeneration in Opuntia ficus-indica (L.) Mill. (Cactaceae). Sci Hortic (Amsterdam). 2006;108: 15–21. doi:10.1016/j.scienta.2005.12.007
22. Arellano-Perusquía A, López-Peralta M, Chablé-Moreno F, Estrada-Luna A. Effect of growth regulators on the organogenesis and multiplication of Ortegocactus macdougalli Alexander . Propag Ornam Plants. 2013;13: 160–167.
23. Vidican I. Studiens on the influence of acid concentration-B indolilbutiric (IBA) on regenerative capacity and organogenesis of explants of Opuntia (Tournef.) Mill. fragilis var. fragilis. J Analele. 2012;19: 312–319.
24. Sriskandarajah S, Prinsen E, Motyka V, Dobrev P, Serek M. Regenerative capacity of cacti Schlumbergera and Rhipsalidopsis in relation to endogenous phytohormones cytokinin oxidase/dehydrogenase, and peroxidase activities. J Plant Growth Regul. 2006;25: 79–88.
25. Rubluo A, Marín-Hernández T, Duval K, Vargas A, Márquez-Guzmán J. Auxin induced morphogenetic responses in long-term in vitro subcultured Mammillaria san-angelensis Sa. Sci Hortic (Amsterdam). 2002;95: 341–349.
26. Clayton PW, Hubstenberger JF, Phillips GC. Micropropagation of members of the Cactaceae Subtribe Cactinae. J Am Soc Hortic Sci. 1990;115: 337–343.
27. Peng H, Zhang J. Plant genomic DNA methylation in response to stresses: Potential applications and challenges in plant breeding. Prog Nat Sci. National Natural Science Foundation of China and Chinese Academy of Sciences; 2009;19: 1037–1045. doi:10.1016/j.pnsc.2008.10.014
28. Neelakandan AK, Wang K. Recent progress in the understanding of tissue culture-induced genome level changes in plants and potential applications. Plant Cell Rep. 2012;31: 597–620. doi:10.1007/s00299-011-1202-z
29. Smulders MJM, de Klerk GJ. Epigenetics in plant tissue culture. Plant Growth Regul. 2011;63: 137–146. doi:10.1007/s10725-010-9531-4
30. Miguel C, Marum L. An epigenetic view of plant cells cultured in vitro: somaclonal variation and beyond. J Exp Bot. 2011;62: 3713–25. doi:10.1093/jxb/err155
31. Briton N, Rose J. The Cactaceae, Hylocereanae Vol II. Dover, New York; 1963.
32. Martínez Ruiz ER. Fenología y desarrollo de pitahaya (Hylocereus undatus Haw. Britt. & Rose) en la región central de Veracruz. Colegio de Postgraduados Montecillo. 2014.
33. Aguirre Z, Kvist LP, Sánchez O. Bosques secos en Ecuador y su diversidad. Botánica Económica los Andes Cent. 2006;12: 162–187.
34. Bauk K, Pérez R, Zeballos S, Peñas L, Flores J, Gurvich D. Are seed mass and seedling size and shape related to altitude? Evidence in Gymnocalycium monvillei (Lem.) Britton & Rose (Cactaceae). Bot ja. 2015;93: 529–533. doi:10.1139/cjb
35. Ofori D, Newton A, Leakey R, Grace J. Vegetative propagation of Milicia excelsa by leaft stem cuttings: effects of auxin concetration, leaf area and rooting medium. For Ecol Manage. 1996;84: 39–48.
36. UNEP-WCMC (Comps.). Index of CITES species. CITES. 2014.
37. Touati A, Lachachi S, Yahia N, Fyad-Lameche F. In vitro variability of morphogenetic responses of tomato (Lycopersicon esculentum Mill.) explants under salt stress and thidiazuron as plant growth regulator. J Appl Biol Sci. 2015;9: 43–49.
38. Rai MK, Kalia RK, Singh R, Gangola MP, Dhawan a. K. Developing stress tolerant plants through in vitro selection—An overview of the recent progress. Environ Exp Bot. 2011;71: 89–98. doi:10.1016/j.envexpbot.2010.10.021
39. Cisneros A, Tel-Zur N. Evaluation of Interspecific-Interploid Hybrids (F1) and Back Crosses (BC1) in Hylocereus Species (Cactaceae). In: Swan A, editor. Meiosis-Molecular Mechanisms and Cytogenetic Diversity. Rijeka, Croatia: In tech; 2012. p. 472.
40. Tel-Zur N, Abbo S, Bar-Zvi D, Mizrahi Y. Genetic relationships among Hylocereus and Selenicereus Vine Cacti (Cactaceae): Evidence from Hybridization and Cytological Studies. Ann Bot. 2004;94: 527–534. doi:10.1093/aob/mch183
41. Perez-Molphe-Balch E, Perez-Reyes ME, Villalobos-Amador E, Meza-Rangel E, Morones L, Lizalde H. Micropropagation of 21 species of mexican cacti by axillary proliferation. Vitr Cell Dev Biol - Plant. 1998;34: 131–135.
42. Santos-Diaz MDS, Mendez-Ontiveros R, Arredondo-Gómez A, Santos-Díaz M de L. In vitro organogenesis of Pelecyphora aselliformis erhenberg (Cactaceae). Vitr Cell Dev Biol - Plant. 2003;39: 480–484.
43. Dávila-Figueroa CA, Rosa-Carrillo M de L, Pérez-Molphe-Balch E. In vitro propagation of eight species or subspecies of Turbinicarpus (Cactaceae). Vitr Cell Dev Biol - Plant. 2005;41: 540–545. doi:10.1079/IVP2005668
44. Rojas-Arechiga M, Vázquez-Yanes C. Cactus seed germination: a review. J Arid Environ. 2000;44: 85–104.
45. Murashige T, Skoog F. A revised medium for rapid growth and bioassays with tobacco. Phy. 1962;15: 473–497.
46. Mohamed-Yasseen Y. Micropropagation of pitaya (Hylocereus undatus Britton et Rose). Vitr Cell Dev Biol - Plant. 2002;38: 427–429. doi:10.1079/IVP2002312
47. Mohamed-Yasseen Y, Barringer S, Splittstoesser W. Rapid propagation of tuna (Opuntia ficus-indica) and plant establishment in soil. Plant Cell Tissue Organ Cult. 1995;42: 117–119.
48. López-Gómez R, Díaz-Pérez J, Flores-Martínez G. Vegetative propagation of three species of Cacti: Pitaya (Stenocereus griseus), Tunillo (Stenocereus stellatus) an jiotilla (Escontria chiotilla). Agrociencia. 2000;34: 363–367.
49. Bhau BS. Regeneration of Coryphantha elephantidens (Lem.) Lem. (Cactaceae) from root explants. Sci Hortic (Amsterdam). 1999;81: 337–344.
50. R Developmente Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-00051-07-0. URL http://www.R-project.org. 2015.
51. Suárez R, Caetano C, Ramírez H, Morales J. Multiplicación de Selenicereus megalanthus (pitahaya amarilla) e Hylocereus polyrhizus (pitahaya roja) vía organogénesis somática. Acta Agronómica. 2014;63: 272–281.
52. Pineda A, Vargas T, García E. Regeneración de Ananas comosus (L.) Merr, ecotipo TABË KÄNÄ, mediante organogénesis indirecta. Bioagro. 2014;26: 135–142.
53. Dávila-Figueroa C, Perez Molphe Balch E. In vitro propagation of Pelecyphora aselliformis ehrenberg and P. strobiliformis werdermann (Cactaceae). Vitr Cell Dev Biol - Plant. 2002;38: 73–78. doi:10.1079/IVP2001248
54. Moebius-Goldammer KG, Mata-Rosas M, Chávez-Avila VM. Organogenesis and somatic embryogenesis in Ariocarpus kotschoubeyanus (Lem.) K. Schum. (Cactaceae), an endemic and endangered Mexican species. Vitr Cell Dev Biol - Plant. 2003;39: 388–393. doi:10.1079/IVP2003427
55. Mosco A. Tissue localization of betacyanins in cactus stems. Rev Mex Biodivers. 2012;83: 413–420.
56. Gibson A, Nobel P. The cactus primer. Harvard Un. Cambridge, Massachusetts.; 1986.
57. Siddiqui MW, Singh JP, Nayyer MA, Barman K, Ahmad MS, Kumar V. 6-Benzylaminopurine affects lipid peroxidation and membrane permeability and thereby preserves curd quality and antioxidants during storage of cauliflower. Acta Physiol Plant. Springer Berlin Heidelberg; 2015;37: 96–104. doi:10.1007/s11738-015-1848-1
58. Wilson-García CY, Zavaleta-Mancera HA, López-Delgado H, Hernández-Garay A. La citocinina BAP retrasa senescencia, aumenta antioxidantes, proteína y crecimiento en el pasto ovillo (Dactylis glomerata L.). Agrociencia. 2008;42: 799–806.
59. Pillay I, Railton I. Complete release of axillary buds from apical donance in htact, light-grown seedlings of Pisum sativum L . following a single application of cytokinin. Plant Physiol. 1983;71: 972–974.
60. Guo B, Abbasi BH, Zeb A, Xu L, Wei Y. Thidiazuron: A multi-dimensional plant growth regulator. African J Biotechnol. 2011;10: 8984–9000. doi:10.5897/AJB11.636
61. Murch S, KrishnaRaj S, Saxena P. Thidiazuron-induced morphogenesis of Regal geranium (Pelargonium domesticum): A potential stress response. Physiol Plant. 2006;101: 183–191.
62. Huetteman C, Preece J. Thidiazuron: a potent cytokinin for woody plant tissue culture. Plant Cell Tissue Organ Cult. 1993;33: 105–119.
63. Ramírez-Mosqueda M a., Iglesias-Andreu LG. Indirect organogenesis and assessment of somaclonal variation in plantlets of Vanilla planifolia Jacks. Plant Cell Tissue Organ Cult. 2015;Publish on. doi:10.1007/s11240-015-0868-2
64. Pant N, Agrawal R, Agrawal S. Mannitol-induced drought stress on calli of Trigonella foenum-graecum L. Var. RMt-303. Indian J Exp Biol. 2014;52: 1128–1137.
65. Nogueira-Nunes E, Bezerra de Sousa AS, Marques de Lucena C, Silva S de M, Paiva de Lucena RF, Belarmino Alves CA, et al. Pitaia (Hylocereus sp.): Uma revisão para o Brasil. Gaia Sci. 2014;8: 90–98.
66. Pérez-Molphe-Balch E, Pérez-Reyes ME, De La Rosa-Carrillo M de L. In Vitro conservation of Turbinicarpus (Cactaceae) under slow growth conditions. BioOne Haseltonia. 2012;17: 51–57.
67. Mauseth JD. Giant shoot apical meristems in cacti have ordinary leaf primordia but altered phyllotaxy and shoot diameter. Ann Bot. 2004;94: 145–53. doi:10.1093/aob/mch121
68. Han H, Zhang S, Sun X. A review on the molecular mechanism of plants rooting modulated by auxin. African J Biotechnol. 2009;8: 348–353.
69. Pierre-Jerome E, Moss BL, Nemhauser JL. Tuning the auxin transcriptional response. J Exp Bot. 2013;64: 2557–63. doi:10.1093/jxb/ert100
70. Frébort I, Kowalska M, Hluska T, Frébortová J, Galuszka P. Evolution of cytokinin biosynthesis and degradation. J Exp Bot. 2011;62: 2431–2452. doi:10.1093/jxb/err004
71. Molinier J, Ries G, Zipfel C, Hohn B. Transgeneration memory of stress in plants. Nature. 2006;442: 1046–1049. doi:10.1038/nature05022
72. Źróbek-Sokolnik A. Temperature stress and responses of plants. In: Ahmad P, Prasad MNV, editors. Environmental adaptations and stress tolerance of Plants in the Era of Climate Change. Springer S. New York, NY: Springer New York; 2012. pp. 113–134. doi:10.1007/978-1-4614-0815-4
73. Schulze J. Improvements in cereal tissue culture by thidiazuron: A review. Fruit, Veg Cereal Sci Biotechnol. 2007;1: 64–79.
74. Sánchez C, Fischer G, Wilson D. Stomatal behaviour in fruits and leaves of the purple passion fruit (Passiflora edulis Sims) and frutis and cladodes of the yellow pitaya (Hylocereus megalanthus (K. Schum. ex Vaupel) Ralf Bauer). Crop Agron. 2013;31: 38–47.
75. Shiskova S, Moreno N, Castillo-Díaz V, Arellano J, Dubrovsky J. Variabilidad genotípica de cactáceas con crecimiento determinado de la raíz en la regeneración de raíces a partir de callos. Zo Áridas. 2006;10: 41–58.
76. Li S-W, Xue L. The interaction between H2O2 and NO, Ca2+, cGMP, and MAPKs during adventitious rooting in mung bean seedlings. Vitr Cell Dev Biol - Plant. 2010;46: 142–148. doi:10.1007/s11627-009-9275-x
77. Li S-W, Xue L, Xu S, Feng H, An L. Mediators, genes and signaling in adventitious rooting. Bot Rev. 2009;75: 230–247. doi:10.1007/s12229-009-9029-9
78. Taiz L, Zeiger E. Plant Cells. Universita. 2006.
79. Valladares F, Gianoli E. Tansley review Ecological limits to plant phenotypic plasticity. New Phytol. 2007;176: 749–763.
80. Poljuha D, Balen B, Bauer A, Ljubesic N, Krsnik-Rasol M. Morphology and ultrastructure of Mammillaria gracillis (Cactaceae) in in vitro culture. Plant Cell Tissue Organ Cult. 2003;75: 117–123.
81. Papafotiou M, Balotis GN, Louka PT, Chronopoulos J. In vitro plant regeneration of Mammillaria elongata normal and cristate forms. Plant Cell Tissue Organ Cult. 2001;65: 163–167.
82. Sultan SE. Phenotypic plasticity for plant development, function and life history. Trends Plant Sci. 2000;5: 537–542. doi:10.1016/S1360-1385(00)01797-0
83. Gratani L. Review Article Plant Phenotypic Plasticity in Response to Environmental Factors. Adv Bot. 2014;Article ID: 1–17. doi:dx.doi.org/10.1155/2014/208747
Recibido: 24 mayo 2017
Aceptado: 25 julio 2017
Máximo Moreira-Palacios1*, Aminael Sánchez-Rodríguez1
1 Universidad Técnica Particular de Loja, Departamento de Ciencias Biológicas. San Cayetano Alto, Apartado Postal 11-01-608, Loja, Ecuador.